The fidelity of chromosome inheritance is of paramount importance to all living organisms. This means that in every cell division one complete copy of the parental genome must be sorted into each of the two daughter cells without any duplication or loss. This is achieved by first the complete and faithful duplication of the genome by the DNA replication machinery. Next the replicated chromosomes need be fully resolved from one another ensuring that no physical connection exists between them. Finally the sister chromosomes need to be segregated to opposite ends of the dividing cell ensuring that when the cell division is completed both daughter cells have a full complement of genetic information.
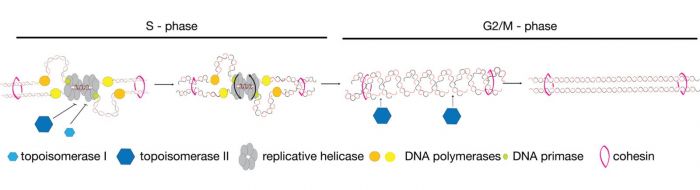
The process of chromosome resolution is essential because the replication of the double helix initially produces sister chromatids with numerous inter-links which initially bind the chromatids together. These would, if left unresolved, prevent segregation of the chromosomes into the daughter cells.
These double stranded interlink between the sister-chromatids or catenanes are mostly generated at the completion of DNA replication. During DNA replication in eukaryotes both topoisomerases I and II (Top2 in yeast) operate ahead of the replisome to relax the supercoils generated by replicative helicase mediated unwinding. In doing so they resolve the majority of links between the parental strands. However, the remaining links between the parental strands are converted into double-stranded DNA catenanes at the termination of DNA replication. Resolving the remaining mitotic catenanes is the essential function of topoisomerase II alone.
The research in the laboratory focuses on two aspects of this process:
- Describing the nature of the replisome during termination phase of DNA replication swivel and the factors that determine when and how replisome swivelling takes place.
- How does the cell ensure that all DNA catenanes or intertwines are resolved before the chromosome segregate the replicated sister chromatids at anaphase.
Central to both these processes is the topology and structure of the DNA double helix. The stress due to DNA topology at termination drives the swiveling of the replisome that occurs at termination resulting in a final replication intermediate of catenated sister chromatids. Conversely during mitosis the sister chromatids are subjected to a dynamic positive supercoiling activity that drives the resolution of catenated sister chromosomes.